利用嗜神经病毒解析小鼠大脑梨状皮层的神经环路毕业论文
2020-04-10 14:47:17
摘 要
梨状皮层是哺乳动物嗅皮层中最大的一块区域,在嗅觉信息的处理过程中起着十分重要的作用,与癫痫的产生也有密切的关系。然而,梨状皮层内的神经元通过十分复杂的兴奋性突触和抑制性突触神经网络产生连接,这给梨状皮层的功能研究带来极大不便。为进一步解析梨状皮层的神经环路,我们利用经基因工程改造的嗜神经病毒rAAV-CaMKⅡ-Cre,rAAV-Ef1α-DIO-EGFP-2a-TK和HSV-ΔTK-LSL-tdTomato,成功标记到了小鼠后梨状皮层中谷氨酸能神经元的顺向单级神经网络。实验结果不仅证实了前人文献中报道过的梨状皮层相关的神经环路,还发现了后梨状皮层中的谷氨酸能神经元的新的投射。本研究为梨状皮层和相关脑区的功能连接研究提供了重要的结构基础。
关键词:嗜神经病毒;后梨状皮层;神经环路;跨突触追踪
Abstract
The piriform cortex (PC), largest area of the mammalian olfactory cortex,plays an important role in epilepsy and odor coding. However, PC neurons are intricately connected through a network of recurrent excitatory and inhibitory synapses, which caused great inconvenience to the relevant research on the function of PC. In order to dissect the neural networks of PC further, we took advantage of gene engineering neurotropic viruses rAAV-CaMKⅡ-Cre, rAAV-Ef1α-DIO-EGFP-2a-TK and HSV-ΔTK-LSL-tdTomato, and successfully mapped the direct output neural networks of glutamatergic neurons in posterior piriform cortex (pPC). The results confirmed the networks relevant to PC which were already known and also showed new projections from glutamatergic neurons in posterior piriform cortex. Our work in this paper mainly prepares for further research on the functional connectivity of PC and relevant brain region.
Key words: neurotropic viruses; posterior piriform cortex; neural network; trans-synaptic tracing
目 录
第1章 绪论 1
1.1梨状皮层简介 1
1.1.1梨状皮层的基本结构 1
1.1.2梨状皮层的输入环路 2
1.1.3梨状皮层的输出环路 4
1.2嗜神经病毒简介 4
1.2.1腺相关病毒 4
1.2.2单纯疱疹病毒 5
1.2.3其他嗜神经病毒 6
1.3研究内容及意义 8
第2章 实验 9
2.1实验材料 9
2.1.1 实验动物 9
2.1.2 病毒 9
2.1.3 主要试剂 9
2.1.4 主要仪器设备 10
2.1.5 实验耗材 10
2.1.6 溶液配制 10
2.2实验方法 11
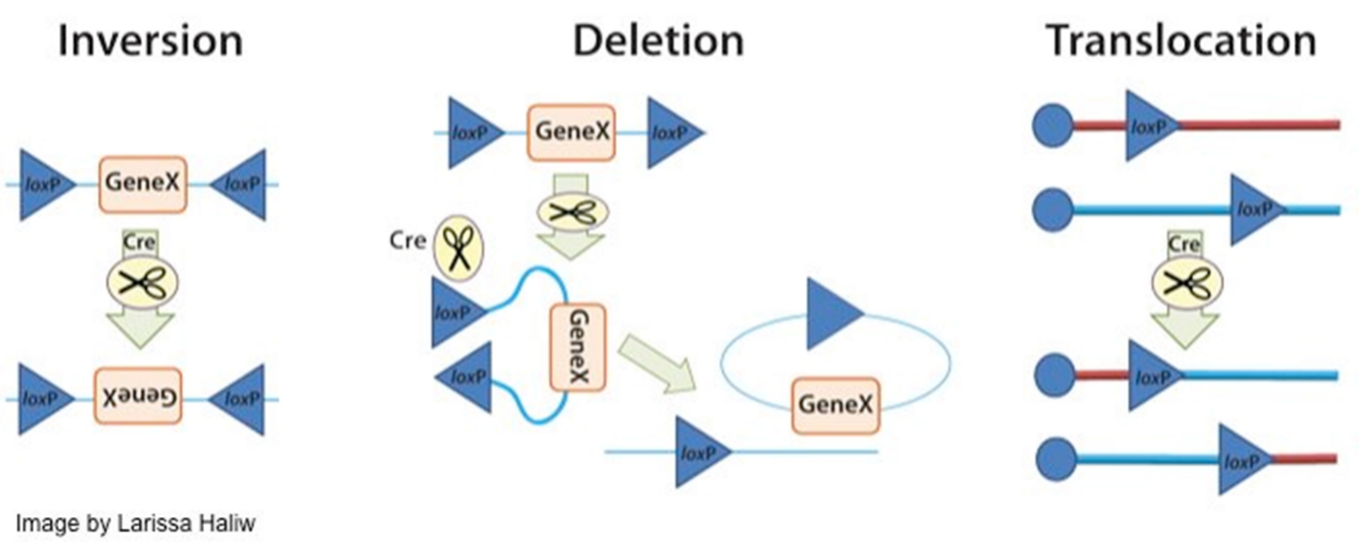
2.2.1顺向单级示踪实验原理 11
2.2.2顺向单级示踪实验方案 14
2.2.3病毒注射 15
2.2.4灌流取样 17
2.2.5样本制备 17
2.2.6显微成像及数据处理 18
第3章 结果 19
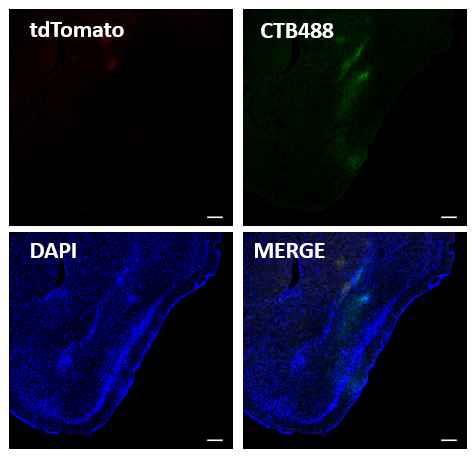
3.1病毒系统验证 19
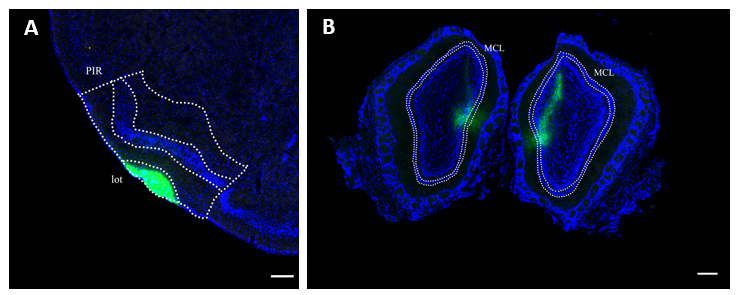
3.2起始细胞的定位 20
3.3后梨状皮层谷氨酸能神经元同侧投射 21
3.4后梨状皮层谷氨酸能神经元对侧投射 24
第4章 总结与展望 27
4.1总结 27
4.2展望 27
参考文献 29
致谢 33
第1章 绪论
1.1梨状皮层简介
梨状皮层(piriform cortex,PC)是动物大脑中的一个脑区,是位于端脑的嗅脑的一部分,与嗅觉功能密切相关。梨状皮层是直接接受来自嗅球的投射输入的最大的皮层区域,且不同于其他感觉皮层的是,PC无需经过丘脑来接收感觉输入,在哺乳动物嗅觉处理进程中有着至关重要的作用,近年来常作为皮层感觉处理的模式环路来进行研究[1]。
1.1.1梨状皮层的基本结构
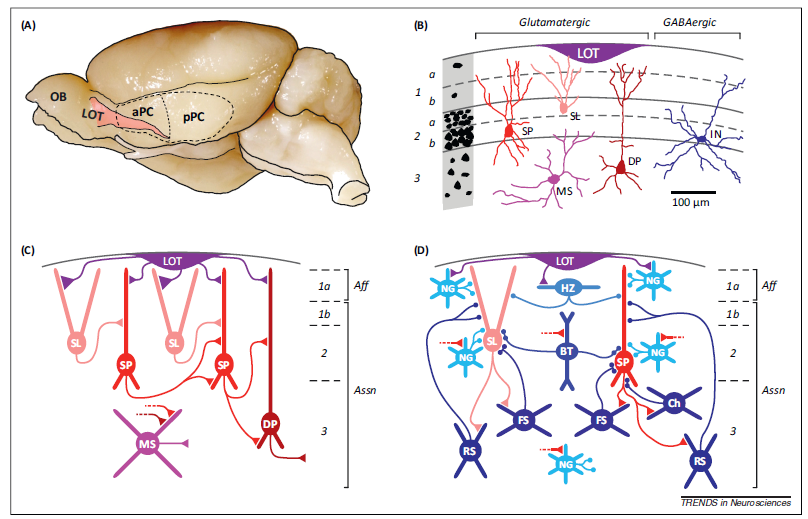
图1.1 梨状皮层的基本结构[1]
在啮齿类动物的脑部结构中,梨状皮层位于大脑表面的腹外侧,与侧嗅束(lateral olfactory tract, LOT)相连,是具有层状结构的旧皮质(如图1.1A)。PC包括神经元稀疏分布的表层(layer1)、包含密集的谷氨酸能神经元的主要输入层(layer2)和包含密度稍低的神经元的深层(layer3)(如图1.1B) [1]。
LOT的输入纤维一般集中于第一层的外侧(layer1a),而来自PC和其他脑区的神经元的联系和联合纤维则分布在layer 1b、layer2和layer3(如图1.1C)[2,3,4]。不同类型的GABA能中间神经元可以为主神经元提供前馈和反馈突触抑制(如图1.1D)。PC也与其他邻近区域存在突触连接,如内梨状核(endopiriform nucleus)、前嗅核(anterior olfactory nucleus)、嗅结节(olfactory tubercle)和皮质杏仁核(cortical amygdala)[2,3]。此外,来自其他脑区的投射输入可以通过释放神经递质来调节PC的活动,如乙酰胆碱和去甲肾上腺素[5,6]。
PC可以简单分为前梨状皮层(anterior piriform cortex, aPC)和后梨状皮层(posterior piriform cortex, pPC)。aPC接受大量来自嗅球的输入和少量联系输入(associational input),而后梨状皮层接受大量的联系输入和少量的嗅球输入[3,7-10]。有研究表明,aPC与外部世界有更强的联系,主要负责气味信息的辨别,而后梨状皮层与气味特性的编码有关[11-16]。
1.1.2梨状皮层的输入环路
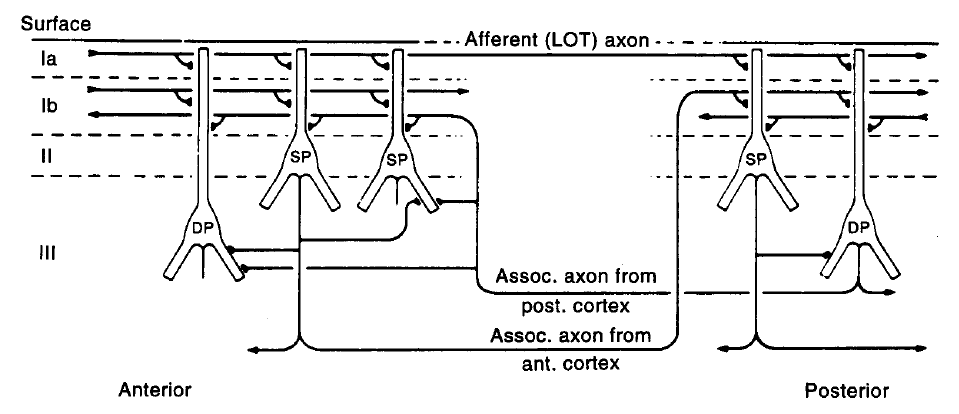
图1.2 梨状皮层的环路基础[17]
PC主要接受来自嗅球僧帽细胞层的输入。来自嗅球带髓鞘的轴突沿着前梨状皮层的表面通过LOT通路到达PC[18]。穿过LOT后,这些轴突将沿着整个PC表面分布,并与分布在layer1a中锥体细胞的树突形成突触连接(如图1.2)。通过电子显微镜,人们发现这些突触是不对称的,表明这些突触行使着兴奋性神经传递的功能。一般认为,在嗅球投射到PC这条通路中,谷氨酸是最主要的神经递质[19]。
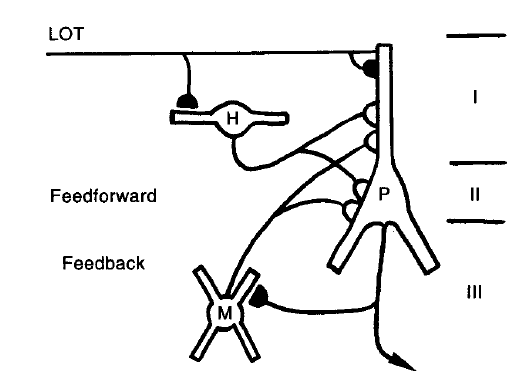
图1.3梨状皮层中的抑制性连接[17]
除了嗅球以外,PC还存在一些自身的突触连接并接受来自基底前脑(basal forebrain)、丘脑(the thalamus)、下丘脑(the hypothalamus)和脑干(the brainstem)的投射。这些自身的输入可以分为兴奋性和抑制性的突触连接(如图1.3)。兴奋性连接的特点是无髓鞘包裹的纤维、来源于PC锥体细胞轴突分支的不对称突触[20,21,22]。这些轴突分支与多极细胞相连或者穿过很大一段距离与layer1b中的锥体细胞的树突进行连接。这些多级细胞和锥体细胞的连接是反馈抑制的结构基础[22]。一些没有被髓鞘包裹的轴突分支穿过Layer3与layer1b中的锥体细胞相连,这些轴突分支被称为联合纤维(association fibers)。PC自身的抑制性输入来源于layer1的水平细胞(horizontal cells)和layer3的多极细胞(multipolar cells),这两种细胞都形成对称的突触结构。水平细胞行使前馈抑制,锥体细胞行使反馈抑制[17]。
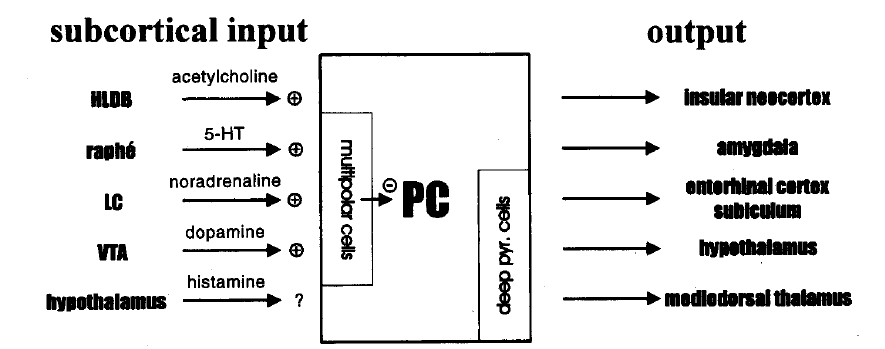
图1.4 梨状皮层的输入和输出[17]
此外,PC还接受一些来自皮层下的输入,这些输入可以调节锥体细胞和中间神经元的活动[17]。有研究表明PC中弥散分布的胆碱能神经元可以降低layer1b层中突触活动的水平,也能引起锥体细胞和中间神经元剧烈去极化之后的再次去极化。胆碱能的调制作用被认为与PC的记忆功能有密切关系[23]。PC也接受来自蓝斑(locus coeruleus, LC)的去甲肾上腺素能的投射和来自腹侧被盖区(ventral tegmental area, VTA)的多巴胺能的投射。
1.1.3梨状皮层的输出环路
PC的输出主要依靠锥体细胞的轴突(如图1.4)。除了梨状皮层内部中间神经元的相互投射以及PC与内梨状核(endopiriform nuclei)之间的相互投射外,梨状皮层的锥体细胞还对同侧的嗅球有很强的投射[24,25]。前梨状皮层可以投射到对侧的梨状皮层[26]。接受梨状皮层投射的非嗅觉系统区域有新皮质岛叶区(insular areas of the neocortex)和杏仁核(amygdala)的各亚区[27-30]。PC内侧部的锥体细胞也可以投到内嗅皮层(entorhinal),下托(subiculum),背内侧丘脑(mediodorsal thalamus),下丘脑(hypothalamus),腹侧壳核(ventral putamen)[31-37]。背内侧丘脑核可以投射到岛叶,岛叶也同时接受PC的直接投射[38]。因此,PC与同侧的嗅觉系统中的核团有紧密的联系,但同时也与新皮质岛叶,边缘系统(limbic system),下丘脑和中脑的运动系统有紧密联系(如图1.4)。
1.2嗜神经病毒简介
1.2.1腺相关病毒
腺相关病毒(adeno-associated virus, AAV)属于细小病毒科(parvoviridae),是无囊膜的单链DNA 病毒,病毒结构为直径20nm的正二十面体(如图1.5A),其基因组大小约为4.7 kb(如图1.5B)。经过改造的rAAV病毒工具已广泛应用于动物水平的基因操作、基因表达和基因治疗研究。AAV是目前为止发现的一类结构最简单的单链DNA缺陷型病毒,通常在辅助病毒的帮助下才能发生毒性感染,产生子代AAV[39]。
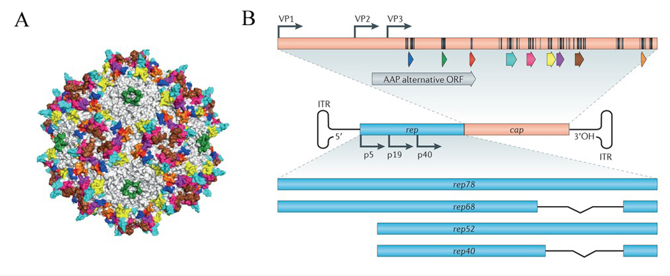
图1.5 A:AAV病毒颗粒示意图;B:AAV病毒基因组结构[39]
由于AAV感染神经系统的免疫原性低、毒性低,且基因表达持久,AAV在神经环路示踪以及相关的功能研究中有极其广泛的应用。
根据其衣壳蛋白的的血清型和相应的受体的区别,AAV对神经系统有不同的感染特性。神经科学中常用的血清型有2、5、8、9、DJ、1/2型,上述血清型的AAV对神经元均有高效感染的能力,其中8、9型比2、5型扩散范围更大;1/2型具有部分逆向转导的特性;5型具有最大的线性DNA组的装载容量[40,41]。在AAV转导神经元的过程中,需要大量的病毒粒子和较长的表达时间,并且需要宿主细胞合成其基因的互补链。scAAV包装了互补DNA,线性DNA组装容量减半,但能介导更强更快的表达[42]。
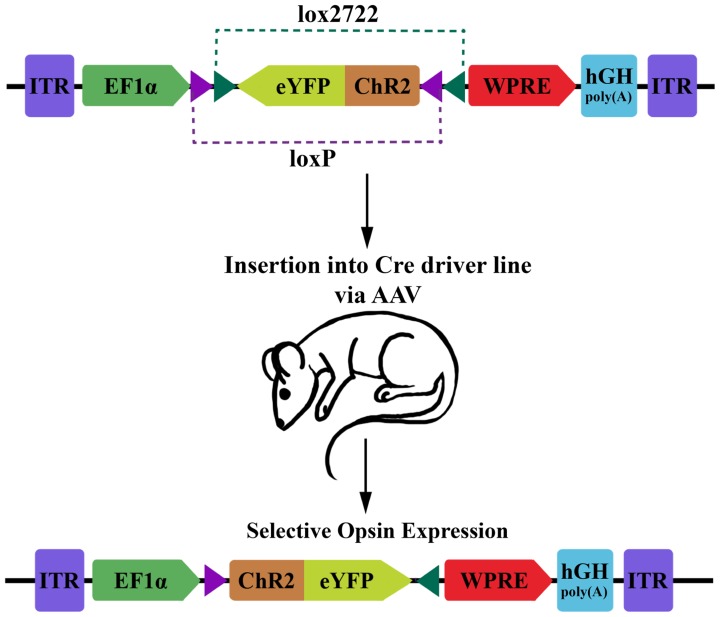
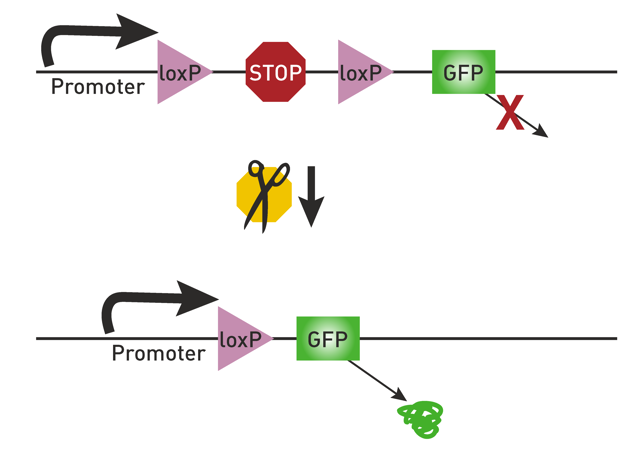
另外,广谱启动子常用于AAV来实现外源基因在宿主细胞内的高效表达,CAG和EF1α较为常用,表达活性也较高;利用特定启动子可以实现对特定类型细胞的感染,例如,利用CaMKⅡ启动子可实现对谷氨酸能神经元的特异性感染;结合cre-loxp等系统可以应用于转基因动物中特定类型细胞的基因表达研究[43]。
1.2.2单纯疱疹病毒
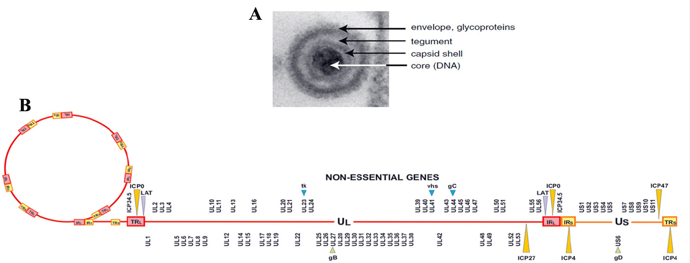
图1.6 A:HSV病毒粒子电镜图; B:HSV基因组构成
(图片来自枢密科技)
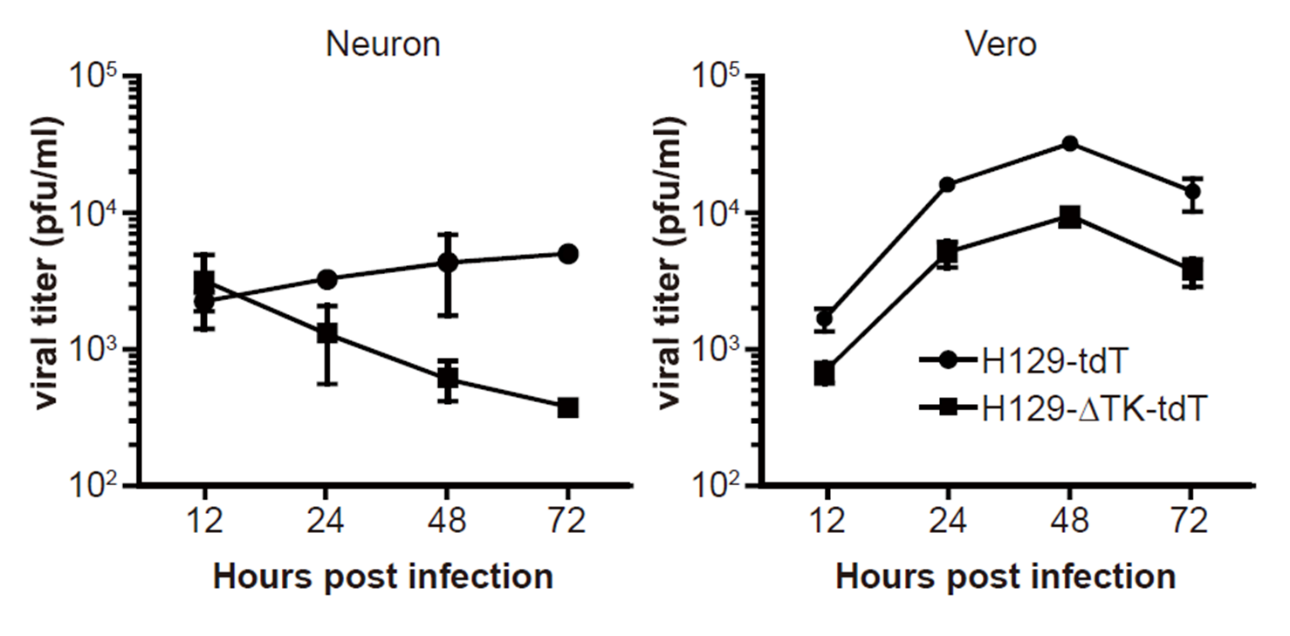
单纯疱疹病毒(Herpes simplex virus,HSV)属于疱疹病毒科、α疱疹病毒亚科,存在HSV-1和HSV-2两种亚型。HSV是线性DNA病毒,其基因组近一半为非必需基因,大小约为152kb。其遗传改造空间大,可容纳大片段外源基因或多个转基因。另外,HSV感染宿主范围广,包括啮齿类和灵长类。HSV-1-129是HSV-1亚型的临床分离株,在外周神经系统(Peripheral nervous system)和中枢神经系统(central nervous system)有严格顺向跨突触的特性。经改造后,装载荧光蛋白的病毒可以高效稳定地表达,从而顺向跨突触标记大脑网络,在神经科学的研究中具有极大的应用价值,适合标记神经的输出环路,是极具潜力的示踪工具。
近年来,有研究者对HSV-1-129病毒株进行了遗传改造。利用同源重组的方法,构建出H129DTK-TT,从而可以通过cre重组酶来控制其荧光表达能力和跨突触能力[44]。利用转基因动物和H129DTK-TT,可以特异性地从嗅觉、视觉感受神经元及浦肯野神经元出发,顺向跨突触追踪全脑地信息输入网络。Dimitrov等[46]利用H129DTK-TT结合Vgatires-Cre/ 转基因小鼠研究了中央杏仁核(centralamygdala, CAmy) 及后外侧下丘脑( posterior lateral hypothalamic area, PLH) GABA 能神经元对蓝斑去甲肾上腺素能神经元的调控网络。其研究结果证实,CAmy 及PLH 中的GABA 能神经元的确对蓝斑核有投射,并与其中去甲肾上腺素能神经元形成突触连接,实现对去甲肾上腺素系统活性的抑制性调节。
1.2.3其他嗜神经病毒
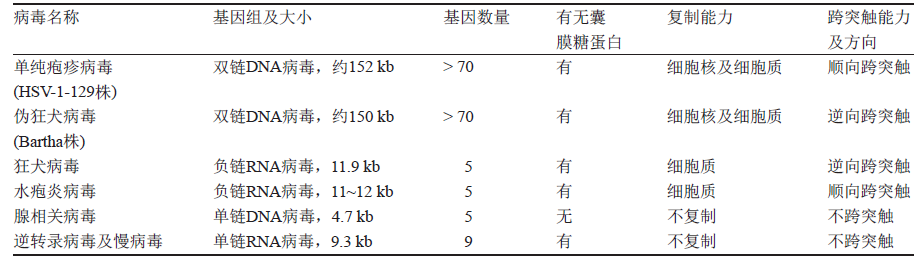
图1.7神经环路研究常用病毒的特性比较[45]
慢病毒载体(Lentiviral vector, LVs)是在HIV-1病毒基础上改造而成的病毒载体系统,它能高效的将目的基因(或RNAi)导入动物和人的原代细胞或细胞系。慢病毒载体基因组是正链RNA,其基因组进入细胞后,在细胞浆中被其自身携带的反转录酶反转为DNA,形成DNA整合前复合体,进入细胞核后,DNA整合到细胞基因组中。整合后的DNA转录mRNA,回到细胞浆中,表达目的蛋白或产生RNAi干扰。
慢病毒载体介导的基因表达或RNAi干扰作用持续且稳定,原因是目的基因整合到宿主细胞基因组中,并随细胞基因组的分裂而分裂。另外,慢病毒载体能有效感染并整合到非分裂细胞中。以上特性使慢病毒载体与其它病毒载体相比,比如不整合的腺病毒载体、整合率低的腺相关病毒载体、只整合分裂细胞的传统逆转录病毒载体,有鲜明的特色。大量文献研究表明,慢病毒载体介导的目的基因长期表达的组织或细胞包括脑、肝脏、肌肉、视网膜、造血干细胞、骨髓间充质干细胞、巨噬细胞等。
慢病毒表达载体删除了HIV病毒绝大多数基因,仅保留了HIV病毒的LTR序列,包装信号,Rev应答原件等。因此免疫原性低,在注射部位无细胞免疫反应,体液免疫反应也较低,不影响病毒载体的第2次注射。
狂犬病毒(rabies virus,RV)属于弹状病毒科(Rhabdoviridae)狂犬病毒属(Lyssavirus)。野生型RV是人畜共患的狂犬病的致病因子,在神经系统中具有传播属性,对人和动物均具有较高致病性。
随着反向遗传学手段的成熟,经过改造的RV,可用于神经回路研究。RV感染中枢系统后,主要标记神经元,几乎不标记胶质细胞;被感染的神经元在一定时间内(7-12天)几乎不发生明显病变及裂解。Wickersham 等[63]基于RV疫苗株Sad B-19感染性克隆构建的复制缺陷型重组RV具有较低的毒性和较高的安全性,可清晰的标记神经元的精细形态,并通过反向互补策略实现了对神经网络联接的逆向跨单级突触追踪。
水泡性口炎病毒(Vesicular stomatitis virus,VSV) 属于弹状病毒科(Rhabdoviridae) 水泡病毒属(Vesiculovirus),是有囊膜的单股负链RNA 病毒。用于顺向跨突触示踪的VSV工具病毒,在大/小鼠动物模型中的神经回路示踪研究中广泛使用。它也能感染多种动物模型,包括鱼类、禽类,及非人灵长类等。其特点是复制及跨突触速度快,外源基因表达量超高,可获得神经元精细形态。
伪狂犬病毒(Pseudorabies virus,PRV)属于疱疹病毒科α-疱疹病毒亚科成员,宿主范围广泛,包括猪、牛、绵羊、山羊、猫、狗等。PRV 疫苗Bartha株具有逆向跨多级突触的特性,可用于解析大脑中枢及外周的神经网络结构。PRV不感染人及灵长类动物,适用于解析啮齿类等动物的神经环路。对于实验人员具有相对较高的安全性,但是仍须在生物安全二级实验室开展工作。
森林脑炎病毒(Semliki Forest virus,SFV)是正链RNA病毒,基因组长约12kb,基因组分为两个不同的区段,5'端2/3的区域编码与RNA合成相关的非结构蛋白,3'端1/3的区域编码病毒结构蛋白,其中衣壳蛋白包裹基因组RNA形成核衣壳,核衣壳外面有脂质双层膜。病毒最外面是跨膜的E1-E2聚体与分泌型的E3形成三聚体,构成刺突蛋白。SFV作为病毒生命周期和病毒性神经病变研究的模型,已被广泛用于相关领域的研究。由于其广泛的宿主范围和高效的复制能力,也被作为一种生产疫苗和抗肿瘤药物的载体,同时也是基因治疗的一个载体工具。
1.3研究内容及意义
综上所述,梨状皮层在嗅觉信息处理的过程中起着十分重要的作用,也与癫痫的产生有很密切的关系。另外,对记忆处理、脑内兴奋传递过程的研究也涉及到梨状皮层这一区域。梨状皮层在哺乳动物大脑中占据的体积大,相关神经网路多而复杂。但是迄今为止,人们还未对梨状皮层进行精细的亚区分类,梨状皮层也存在许多上游和下游的环路尚未阐明,这些都给梨状皮层更深一步的研究带来不便,所以解析梨状皮层的神经网络具有很大的研究意义。
以上是毕业论文大纲或资料介绍,该课题完整毕业论文、开题报告、任务书、程序设计、图纸设计等资料请添加微信获取,微信号:bysjorg。
相关图片展示: